 2018, Vol. 25
2018, Vol. 25 

2. 广西友谊关森林生态系统国家定位观测研究站,广西凭祥 532699


2. Guangxi Youyiguan Forest Ecosystem National Research Station, Pingxiang, Guangxi, 532699, China
【研究意义】喀斯特季节性雨林是具有地带性植被特征的地域性植被,孕育着丰富的生物多样性,发挥着物种保存、防止石漠化、碳库等重要的生态系统服务功能,是喀斯特地区植被管理和重建的重要参照系,研究其主要种群的生态位特征,有助于了解群落内各种群对资源的利用状况及其生态适应性,对生物多样性的保护及退化生态系统的植被恢复具有重要意义。【前人研究进展】生态位一词是Grinnen于1971年首先提出并使用,如今已经成为生态学的核心概念之一,也是生态学研究的热点问题[1]。生态位特征能反映种群对所处生境的适应能力及其对环境资源的利用能力,是评价物种种间关系、种内关系及种群在群落中所处地位和所起作用的一个重要指标,被广泛应用于生物多样性形成及其维持机制、群落结构与演替以及森林资源保护与利用等方面的研究[2-6]。相关学者对热带[1]、亚热带[5]和温带森林[6]植物种群生态位开展了大量研究,为各地区的森林植被管理、生物资源保护及可持续利用提供了基础性资料和理论指导[7]。喀斯特季节性雨林主要分布于广西西南部的喀斯特峰丛洼地地区,由于特殊的岩溶地质背景,其群落性质、区系组成、森林生态系统对环境的适应等与同一地区的非喀斯特森林明显不同[8]。喀斯特季节性雨林地区从洼地到峰丛顶部高差一般只有200~300 m,但水热条件差异很大,各物种对生境的偏好明显不同,植被垂直变化也很明显:洼地及其边缘,主要分布喜湿耐荫植物,植物的花大多是鲜艳的大型花,倾向于通过萌生方式进行更新;坡地中部,主要分布旱生性较强的植物,植物的花一般是不鲜艳的中小型花,主要通过种子进行更新;山顶及其周围则以山顶矮林为主,植物的花大多数是近白色的小型花,倾向于通过萌生方式进行更新[9-12],不同生境代表性植物的水分利用效率也明显不同(黄甫昭,待发表材料)。这些研究表明喀斯特季节性雨林不同物种的生境偏好、功能性状、更新方式及水分利用策略明显不同,生态位分化明显。【本研究切入点】生态位特征主要包括生态位宽度和生态位重叠,其能定量地评价植物资源利用能力和种间关联性[13]。目前,王斌等[14]、黄甫昭等[15]对喀斯特季节性雨林的物种组成及格局、群落结构及其更新动态进行了相关研究,但缺乏应用生态位理论探讨主要种群对资源利用状况及种群间的竞争共存规律的相关研究。【拟解决的关键问题】本研究以弄岗15 hm2喀斯特季节性雨林监测样地的375个20 m×20 m样方作为作为不同的资源位,以物种重要值作为生态位计测的状态指标,计算分析样地内优势种群的生态位宽度和生态位重叠度,探讨喀斯特季节性雨林优势种群对资源利用的相互关系以及种间竞争机制和规律,为喀斯特季节性雨林生物多样性保护与可持续利用及岩溶石漠化地区植被恢复提供参考。
1 材料与方法 1.1 研究区域概况研究区域位于广西西南部弄岗国家级自然保护区(106°42′28″~107°04′54″E,22°13′56″~22°33′09″N),保护区呈西北-东南长条状分布,总面积10 080 hm2。该区属热带季风气候区,每年有7个月平均气温达22℃,最冷月平均气温13℃,年最高气温37~39℃,≥10℃的积温为7 344~7 930℃,无霜期可达351 d以上。年平均降水量为1 150~1 550 mm。土壤主要有黑色石灰土和棕色石灰土[14]。
保护区内植物资源极为丰富,有蕨类植物和种子植物172科709属1 454种,其中,有包括金花茶(Camellia petelotii)、蚬木(Excentrodendron tonkinense)、金丝李(Garcinia paucinervis)等国家珍稀濒危植物33种,广西重点保护野生植物73种,广西特有植物101种,喀斯特特有植物278种。被子植物以大戟科、茜草科、蝶形花科、禾本科、菊科等泛热带性质科为优势[16-17]。
该区属于喀斯特峰丛-洼地地貌,从洼地到峰丛顶部高差一般只有200~300 m,但生境异质性强,水热条件差异大,植被垂直变化明显。从洼地到峰丛顶部,包含了3种主要生境类型:(1)洼地及其边缘,主要分布喜湿耐荫的植物种类,如海南风吹楠(Horsfieldia hainanensis)、东京桐(Deutzianthus tonkinensis)、中国无忧花(Saraca dives)等;(2)坡地中部,主要分布有蚬木、肥牛树(Cephalomappa sinensis)、闭花木(Cleistanthus sumatranus)等旱生性较强的植物种类;(3)山顶及其周围,主要以细叶谷木(Memecylon scutellatum)、毛叶铁榄(Sinosideroxylon pedunculatum)、米念芭(Tirpitzia ovoidea)为主的山顶矮林[18]。
1.2 样地设置与数据收集弄岗样地位于弄岗保护区弄岗片核心区弄姆皇,地理位置22°25′ N、106°57′ E,样地东西长500 m,南北宽300 m,包含一条山脊和一个较完整的洼地,海拔范围180~370 m[15],坡度范围3.7~78.9°(图 1)。依照CTFS全球森林生物多样性监测规范[19],整个样地用全站仪划分为375个20 m×20 m的样方。调查时,用铝牌标记并测量样方内所有胸径≥1 cm的木本植物,记录其种名、胸径、坐标及生长状况。样地于2011年完成第一次调查工作,获得样地第一次植被及地形因子数据,样地内有监测树种223种,隶属于56科157属,独立个体总数为68 010株[14]。

|
图 1 弄岗样地的三维地形 Fig.1 Topography of the Nonggang plot |
根据样方调查数据,分别计算223个物种在整个样地和各个样方的重要值,选择在整个样地重要值排序前30的物种计算其种群生态位特征。重要值的计算公式如下:
Ⅳ样地=(相对密度+相对优势度+相对频度)/3,
Ⅳ样方=(相对密度+相对优势度)/2,
式中,Ⅳ样地是指物种在整个样地的重要值,Ⅳ样方是指物种在各个样方的重要值,相对密度(%)=100×某个种的株数/所有种的总株数,相对优势度(%)=100×某个种的胸高断面积/所有种的胸高断面积之和,相对频度(%)=100×某个种的频度/所有种的频度之和[20]。
生态位宽度,采用Levins(1968)提出的公式[21],即
| $ {B_i} = \frac{1}{{\sum\limits_{j = 1}^r {{{\left( {{p_{ij}}} \right)}^2}} }}, $ |
式中,Bi为种i的生态位宽度,Pij为种i在j资源位的重要值比例,Pij=nij/Ni,其中Ni=∑nij,nij为种i在j资源位(j样方)的重要值,r为资源位数。
生态位重叠度,采用Pianka生态位重叠公式[22],即
| $ NO = \sum\limits_{j = 1}^r {{n_{ij}} \cdot {n_{kj}}} /\sqrt {{{\left( {\sum\limits_{j = 1}^r {{n_{ij}}} } \right)}^2} \cdot {{\left( {\sum\limits_{j = 1}^r {{n_{kj}}} } \right)}^2}} , $ |
式中,NO为生态位重叠值,nij和nkj为种i和k在j资源位上的重要值。
数据计算采用R3.5.1软件进行,生态位宽度和生态位重叠分别采用种间联结分析程序包spaa中的niche.width()、niche.overlap()函数计算[23]。
2 结果与分析 2.1 主要种群重要值与生态位宽度重要值表明各个种群在群落中的地位和作用,样地主要物种的重要值见表 1。生态位宽度反映植物种群对资源的利用状况,也反映种群在群落中的地位和分布的均匀程度,种群生态位宽度越大,对环境的适应能力越强,对资源的利用更充分[24]。因此,筛选植物对石漠化地区进行植被修复时,生态位宽度可以作为评价指标之一。由表 1可知,喀斯特季节性雨林生态位宽度排序靠前的种群为苹婆、广西牡荆、茎花山柚、假玉桂和日本五月茶,Levins生态位宽度分别为207.04,195.01,176.88,142.05和126.23;生态位宽度排序靠后的是三角车、肥牛树、对叶榕、米扬噎和中国无忧花,分别为37.95,35.79,33.83,25.87和15.76;重要值排序靠前的闭花木、蚬木、海南椴和劲直刺桐的生态位宽度居于中间位置。Pearson相关分析表明,各种群生态位宽度与该种群在整个样地的重要值显著相关(R=0.644、P < 0.001)。苹婆、广西牡荆和假玉桂除了山顶其他地形均有分布,日本五月茶山腰以下普遍分布,茎花山柚也是除了山顶和谷底其他地方广泛分布[25],生态位都较宽,说明这些种的生态幅较宽,各类生境都能适应,对环境变化不是太敏感,在进行植被修复时应优先考虑这些植物种类。闭花木、蚬木和海南椴等是整个样地重要值排序靠前的种群,但它们对环境的要求特殊,分布范围仅局限于山腰的特定地带[25],生态位并不宽。闭花木、蚬木、海南椴等重要值虽大,但是生态位不是特别宽,生态幅和生态适应性并不是特别强,如若将它们作为植被恢复树种,只适宜将其种植于山坡中部特定地带。而对叶榕、米扬噎和中国无忧花等对生境有特殊偏好的物种,主要在特定区域集群分布,生态幅较窄,生态位宽度很小,对环境变化特别敏感,不建议选做植被恢复树种。
| 表 1 弄岗15 hm2喀斯特季节性雨林样地优势种群重要值与生态位宽度 Table 1 The important value and niche breadth of dominant populations in the 15 hm2 karst seasonal rain forest plot |
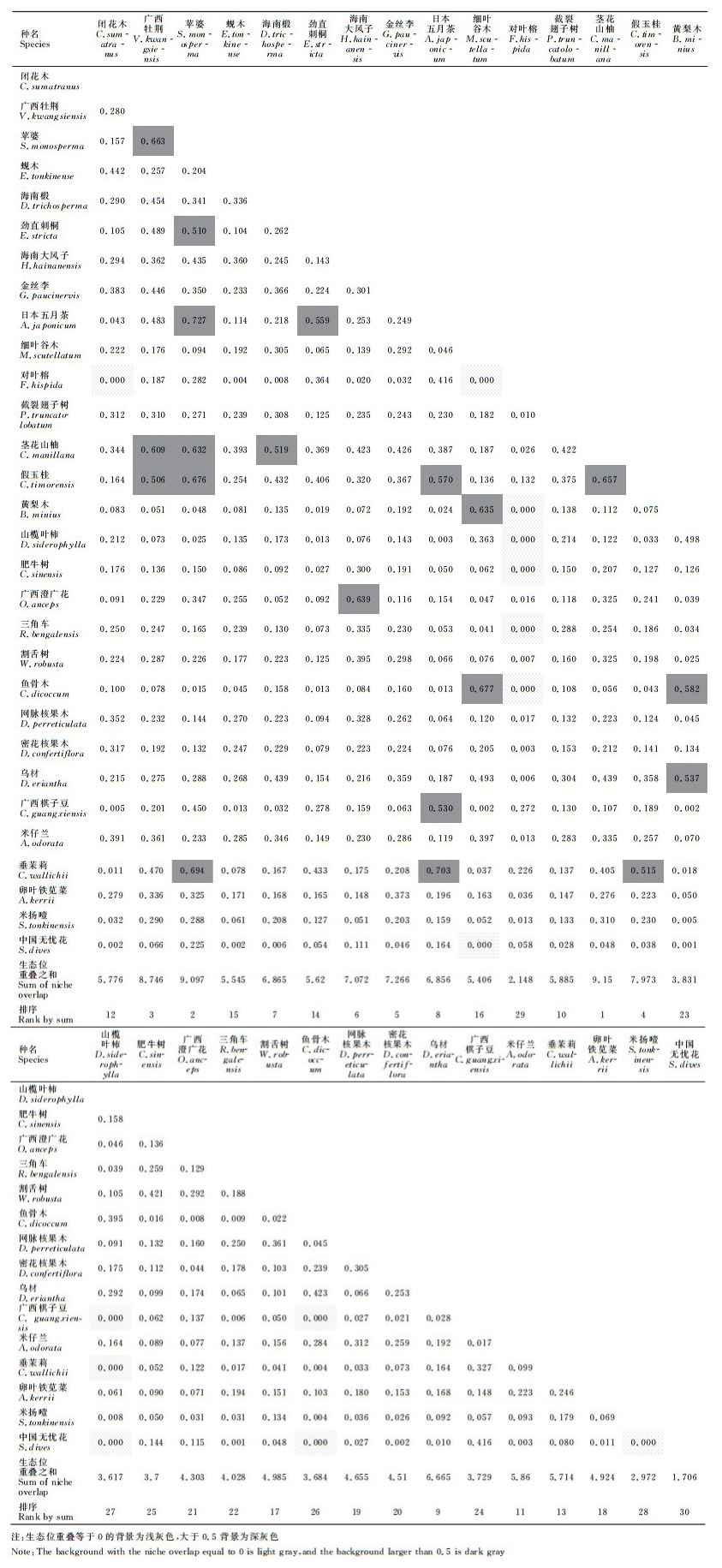
生态位重叠是两个物种在生态因子联系上具相似性,当两个物种利用或共同占有某一资源因素时,就会有生态位重叠。生态位重叠较大的种群生态特性相似,或者对生境因子有互补性的要求,生态位重叠使物种之间发生联系,利用生态位重叠值的大小可以解释物种间的竞争或共存关系[26-27]。弄岗喀斯特季节性雨林15 hm2样地,30个优势种群共组成435个种对关系(表 2),其中,生态位重叠值等于1、完全重叠的种对有0对;生态位重叠值大于或等于0.5的种对有20对,分别为广西牡荆×苹婆(0.663)、广西牡荆×茎花山柚(0.609)、广西牡荆×假玉桂(0.506)、苹婆×劲直刺桐(0.510)、苹婆×日本五月茶(0.727)、苹婆×假玉桂(0.632)、苹婆×茎花山柚(0.676)、苹婆×垂茉莉(0.694)、海南椴×茎花山柚(0.519)、劲直刺桐×日本五月茶(0.559)、海南大风子×广西澄广花(0.639)、日本五月茶×假玉桂(0.570)、日本五月茶×广西棋子豆(0.530)、日本五月茶×垂茉莉(0.703)、细叶谷木×黄梨木(0.635)、细叶谷木×鱼骨木(0.677)、茎花山柚×假玉桂(0.657)、假玉桂×垂茉莉(0.515)、黄梨木×鱼骨木(0.582)和黄梨木×乌材(0.537),占总种对数的4.6%。生态位重叠值大于0小于0.5的种对有401对,占92.2%;生态位重叠度等于0,不发生重叠的种对有14对,占种对数3.2%,分别为闭花木×对叶榕、细叶谷木×对叶榕、细叶谷木×中国无忧花、对叶榕×黄梨木、对叶榕×山榄叶柿、对叶榕×肥牛树、对叶榕×三角车、对叶榕×鱼骨木、山榄叶柿×广西棋子豆、山榄叶柿×垂茉莉、山榄叶柿×中国无忧花、鱼骨木×广西棋子豆、鱼骨木×中国无忧花和米扬噎×中国无忧花。可见,喀斯特季节性雨林主要种群之间的生态位重叠程度普遍较低,群落主要优势种群之间竞争不激烈,群落稳定。各种群与其他种群生态重叠度之和与重要值、生态位宽度的相关性分析表明,与其他种群生态重叠度之和与该种重要值显著相关(R=0.494、P=0.006)、与该种群的生态位宽度也显著相关(R=0.951、P < 0.001)。
| 表 2 弄岗15 hm2喀斯特季节性雨林样地优势种群生态位重叠 Table 2 Niche overlap of of dominant populations in the 15 hm2 karst seasonal rain forest plot |
石漠化是特殊的岩溶作用过程叠加垦荒、砍树、烧山、放牧等人类活动而造成的,是自然因素与人为因素共同作用的结果,石漠化综合治理的首要任务就是植被修复[28]。石漠化地区植被恢复一般都要经历草本群落、灌草群落、灌木灌丛、灌乔过渡和常绿落叶阔叶混交林各个阶段[29]。植被恢复不同阶段,群落环境不同,植物对群落资源的利用和适应能力不同,种间关系也有所不同。而种群生态位特征可以量化评价种群对资源的利用和环境的适应能力及种间竞争关系,为植被恢复设计时物种筛选和配置提供科学依据。但本研究仅对北热带喀斯特地区顶级群落喀斯特季节雨林进行生态位分析,要建立完整的石漠化地区植被恢复参照系,应在不同岩溶气候带建立不同演替阶段的天然植被监测样地,分析其群落环境条件、结构及种间关系等,为不同气候带不同恢复阶段的植被恢复设计提供参考。
当要在某个地区针对某个恢复阶段组建一种恢复模式时,可以先在该演替阶段的天然群落中分析群落环境、树种的生态位特征,选择生态位宽度大的物种作为建群种,以与其它种生态位重叠度小的物种作为伴生种,将不同的种群组配在群落的相应空间位置,合理利用环境资源,使群落处于一种互补互利的协调关系。如果天然群落中各种群的生态位特征不清楚的情况下,盲目将一些生态位重叠很大的种群组合在一起,可能加剧种间竞争,降低森林生态系统中生态位的多样性,使生态系统的稳定性变低[30]。在对一些结构单一的群落进行优化时,我们也要应充分考虑到种群的生态特征,避免引入种与原有种之间产生较大的生态位重叠,防止种群间出现激烈竞争,并通过适当的人为干扰,建立引入种的最适生长环境,使其获得生长优势,使各种群均能有效地利用资源,提高群落的初级生长力[31]。总之,在进行植被修复时,我们应该既要考虑到种群间的竞争排斥原理,尽量不要将生态位重叠很大的物种搭配在一起,同时也要考虑到生态位宽度对植物生存的影响,不要选择对生境有特殊要求、生态位很窄的物种作为先锋种。
3 讨论重要值和生态位宽度是衡量种群在群落中的地位和作用的重要指标,重要值反映的是植物种在群落中的优势度,生态位宽度则表示不同物种对环境资源的利用状况及生态适应幅度等生态位特征,大量研究表明,重要值越大,生态位宽度也越大[24, 32-33]。本研究中,重要值排序第2、第3的广西牡荆、苹婆,其生态位宽度排序第1和第2;重要值最大的闭花木,其生态位宽度排第8;重要值排第4的蚬木,生态位宽度排第15;重要值排在最后两位的米扬噎和中国无忧花,生态位宽度依然排在最后两位。相关分析也表明,种群生态位宽度和重要值具有显著的相关性,但相关系数不是特别高,种群生态位宽度和重要值排序不完全一致。这是因为重要值并不是影响生态位宽度的唯一因素,分布频度也是影响物种生态位宽度的重要因素,分布频度越大,生态位宽度越大[34]。茎花山柚、假玉桂、日本五月茶和金丝李等种群虽然重要值不是特别高,但其环境适应能力较强,在资源位中出现的次数多,分布范围较广,它们的生态位宽度也较大。而像蚬木、对叶榕、中国无忧花,它们对生境有特殊偏好,蚬木偏好干热山腰,对叶榕、中国无忧花偏好湿润谷底,它们在这个局部地带形成优势种,重要值比较大,但在其他生境很少有分布,分布频度并不高,因此,它们的生态位宽度并不宽。可见,重要值与生态位宽度之间不是绝对的线性关系,群落优势种不一定是群落内生态位宽度最大种群,种群生态位宽度受重要值和分布频度等的综合影响。
生态位宽度是一个种群所利用的各种资源的总和,是植物种群对资源利用状况的度量,能反映物种对所属环境的适应状况[35]。植物群落建群种在营造群落内部生境和决定群落内物种构成等方面起主导作用,其生存和适应力都较强,因此建群种的生态位宽度通常比较宽[36]。刘润红等[24]研究漓江河岸带枫杨群落主要木本植物种群生态位特征发现,该群落的建群种枫杨在所有资源位上都有分布,重要值最大,在群落中处于绝对的优势地位,其生态位宽度最大。本研究发现,蚬木是乔木层优势种,是喀斯特季节性雨林主要的建群种,但其主要分布在山坡中部,分布范围不广,生态位宽度只是居于中间位置。可能是因为弄岗样地从谷底到山顶生境异质性很高,水热条件差别很大[15],蚬木对山坡中部生境具有很强的适应性,存在大量高大个体,但在山顶和谷底等生境却鲜有分布,所以蚬木的重要值虽大,但其生态位宽度并不宽。亚乔木层优势种闭花木也具有蚬木相似的特征,重要值虽大,但生态位宽度并不大,也是因为其只分布山坡中上部,其他生境少有分布。
生态位宽度大小可以很好地反映植物种群利用资源和对环境的适应能力,可以依据物种的生态位宽度的大小来选择物种进行植被恢复[37]。喀斯特石漠化地区自然环境恶劣和水土资源匮乏,那些生态位宽度大,资源利用能力强、环境适应性好、竞争能力强的树种应被优先考虑用于植被恢复。苹婆、金丝李、茎花山柚、日本五月茶、广西牡荆和假玉桂是亚乔木层优势种,广泛分布于喀斯特峰丛洼地各种生境类型,是喀斯特季节性雨林生态位宽度比较大的种群,表明它们能充分利用各种资源,对喀斯特环境具有很强的适应性。其中,苹婆在广西热带和南亚热带石山或土山的谷地还是在山坡上都能生长良好,生态适应性广,其种仁风味像板栗,可以煲汤或作烹饪配菜等,在广西石灰岩山区有“木本粮食”之称[38],是兼备经济、生态效益的经济林木,是热带、南亚热带喀斯特地区极具开发潜力的木本粮食植物,在岩溶石漠化地区进行植被恢复时应优先考虑。金丝李木质坚硬、耐腐且抗虫蛀,主要分布于广西西部、南部和云南东南部坡度较陡的喀斯特山地上,是喀斯特石山特有树种,属于中国珍稀濒危保护植物和国家二级保护植物[39]。生态位宽度分析表明,金丝李生态位较宽,能充分利用喀斯特季节性雨林的资源,对其喀斯特生境有较高的适应能力,但金丝李结实的植株极少,果实易被猴子采食且种子寿命短,种子萌发率低,其种苗资源非常有限[40-41],如果能突破繁育技术和苗木资源限制,金丝李将成为北热带和南亚热带石山地区植被恢复的良好材料。茎花山柚是一种石灰岩山地特有植物,对喀斯特季节雨林也具有很强的生态适应性,生态位较宽,在云南、贵州的喀斯特石山地区也有分布,且该种可作为蔬菜食用,具有一定的经济价值,但目前尚未有人针对该种开展相关研究,能否在岩溶石漠化地区推广种植有待进一步研究。日本五月茶、广西牡荆和假玉桂的生态位都较宽,对喀斯特生境适应性较强,但目前尚未发现它们的经济价值,可以做石漠化地区植被恢复的备选树种。
中国无忧花和对叶榕等只分布于谷底,米扬噎等只在山腰中下部局部地区集群分布,表明它们对喀斯特季节性雨林环境的适应范围较窄,对资源的利用能力也较弱,导致它们的生态位宽度小。一般认为生态位宽度较小的物种,对环境资源的利用和适应能力较差,趋向于特种化,它们对环境变化比较敏感,在群落的发展过程中存在被淘汰的危险[24]。所以在对喀斯特季节性雨林进行管护时,应避免对这些种群所在的生境进行干扰。
生态位重叠是评价物种对环境资源利用能力差异性的重要指标,当两个物种共同利用同一资源或共同占有某一资源位时,就会出现生态位重叠[42]。本研究没有发现生态位完全重叠的种对,与生态位完全重叠的物种不能共存理论一致,绝大多数种对的生态位重叠度介于0与0.5之间,优势种群对资源的共享趋势较为明显,群落相对稳定,同时也有少数种对生态位重叠度为0,生态位分化明显。大量研究表明,生态位宽度较大的种群,其资源利用能力较强,分布较广,因而与其他物种间的生态位重叠值也较大[43]。本研究也同样发现,生态位重叠度大于0.5的种对主要是生态位宽度排序靠前的苹婆、广西牡荆、茎花山柚、假玉桂和日本五月茶之间,或者有一个种属于它们的种对。而生态位重叠度为0的种对则主要是生态位宽度排序靠后的对叶榕、广西棋子豆、中国无忧花、肥牛树、三角车、山榄叶柿、鱼骨木和细叶谷木之间。
生态位重叠值越大说明物种对资源的需求越相似,物种之间可能存在激烈竞争[13, 44]。所以在植被恢复的物种选择和配置上,应当既考虑植物种群的生态位宽度大小,又考虑种群间生态位重叠程度[37]。在进行植被恢复时,首先应选择生态位宽度大的乡土物种作为先锋种,以与其生态位重叠小的种群为伴生种,不能将生态位重叠度很大的物种配置在一起,以减少种间竞争,同时也不能将生态位重叠度非常小或者等于0的物种配置在一起,有利于形成稳定的群落,最终达到植被恢复的目的。
4 结论喀斯特季节性雨林中适应性强、分布范围广的种群,生态位宽度较大,且绝大部分种对的生态位重叠度较小,群落主要优势种群之间竞争不激烈,群落稳定。苹婆、金丝李和茎花山柚等生态位宽度比较大,资源利用能力和环境适应能力较强、而且兼具生态和经济价值,在岩溶石漠化地区进行植被恢复时应作为优先考虑物种。要建立完整的石漠化地区植被恢复参照系,应在不同岩溶气候带建立不同演替阶段的天然植被监测样地,分析其群落环境条件、结构及种间关系等,为不同气候带不同恢复阶段的植被恢复设计提供参考。在针对某个地区某个恢复阶段进行植被恢复设计时,应参照该地区这个恢复阶段的天然群落,选择生态位宽度大的物种作为先锋种,选择生态位重叠度较小的物种作为伴生种。
| [1] |
陈瑞国, 范少辉, 刘广路, 等. 海南岛次生低地雨林棕榈藤伴生群落优势种生态位研究[J]. 西北植物学报, 2017, 37(6): 1226-1233. CHEN R G, FAN S H, LIU G L, et al. Niche characteristics of dominant species of rattan accompanying communityin secondary lowland rain forest in Hainan Island, China[J]. Acta Botanica Boreali-Occidentalia Sinica, 2017, 37(6): 1226-1233. |
| [2] |
杨利民, 周广胜, 王国宏. 草地群落物种多样性维持机制的研究Ⅱ物种实现生态位[J]. 植物生态学报, 2001, 25(5): 634-638. YANG L M, ZHOU G S, WANG G H. Investigating realized niches as a mechanism of species diversity maintenance in a species-rich grassland community[J]. Acta Phytoecologica Sinica, 2001, 25(5): 634-638. DOI:10.3321/j.issn:1005-264X.2001.05.020 |
| [3] |
苏志尧, 吴大荣, 陈北光. 粤北天然林优势种群生态位研究[J]. 应用生态学报, 2003, 14(1): 25-29. SU Z Y, WU D R, CHEN B G. Niche characteristics of dominant populations in natural forest in North Guangdong[J]. Chinese Journal of Applied Ecology, 2003, 14(1): 25-29. DOI:10.3321/j.issn:1001-9332.2003.01.006 |
| [4] |
史小华, 许晓波, 张文辉. 秦岭冷杉群落主要种群生态位研究[J]. 植物研究, 2007, 27(3): 345-349. SHI X H, XU X B, ZHANG W H. Study on the niche of the main populations in Abies chensiens community[J]. Bulletin of Botanical Research, 2007, 27(3): 345-349. |
| [5] |
胡正华, 钱海源, 于明坚. 古田山国家级自然保护区甜槠林优势种群生态位[J]. 生态学报, 2009, 29(7): 3670-3677. HU Z H, QIAN H Y, YU M J. The niche of dominant species populations in Castanopsis eyrei forest in Gutian Mountain National Nature Reserve[J]. Acta Ecologica Sinica, 2009, 29(7): 3670-3677. DOI:10.3321/j.issn:1000-0933.2009.07.027 |
| [6] |
刘巍, 曹伟. 长白山云冷杉群落主要种群生态位特征[J]. 生态学杂志, 2011, 30(8): 1766-1774. LIU W, CAO W. Niche characteristics of main plant species in spruce-fir forests in Changbai Mountains[J]. Chinese Journal of Ecology, 2011, 30(8): 1766-1774. |
| [7] |
秦随涛, 龙翠玲, 吴邦利. 茂兰喀斯特森林优势灌木种群的生态位研究[J]. 林业资源管理, 2018(2): 58-64. QIN S T, LONG C L, WU B L. Study on niche of dominant shrub populations in karst forest in Maolan Nature Reserve, Guizhou Province[J]. Forest Resources Mangement, 2018(2): 58-64. |
| [8] |
胡舜士, 王献溥. 广西石灰岩地区季节性雨林的群落学特点[J]. 东北林学院学报, 1980(4): 11-26. HU S S, WANG X P. The phytocoenological features of seasonal rain forest of limestone region in Guangxi[J]. Journal of North-Eastern Forestry Institute, 1980(4): 11-26. |
| [9] |
苏宗明, 赵天林, 黄庆昌, 等. 弄岗自然保护区植被调查报告[J]. 广西植物, 1988, 8(增刊1): 188-214. SU Z M, ZHAO T L, HUANG Q C, et al. The vegetation of Longgang Natural Reserve in Guangxi[J]. Guihaia, 1988, 8(Suppl.1): 188-214. |
| [10] |
郭屹立, 王斌, 向悟生, 等. 弄岗北热带喀斯特季节性雨林15 hm2样地木本植物萌生特征[J]. 生态学杂志, 2015, 34(4): 955-961. GUO Y L, WANG B, XIANG W S, et al. Sprouting characteristics of tree species in the 15 hm2 plot of northern tropical seasonal rain forest in Nonggang, Guangxi, Southern China[J]. Chinese Journal of Ecology, 2015, 34(4): 955-961. |
| [11] |
蒋裕良, 白坤栋, 郭屹立, 等. 北热带喀斯特森林木本植物花性状及其生境分异[J]. 生物多样性, 2016, 24(2): 148-156. JIANG Y L, BAI K D, GUO Y L, et al. Floral traits of woody plants and their habitat differentiations in a northern tropical karst forest[J]. Biodiversity Science, 2016, 24(2): 148-156. |
| [12] |
黄甫昭, 丁涛, 李先琨, 等. 弄岗喀斯特季节性雨林不同群丛物种多样性随的海拔变化[J]. 生态学报, 2016, 36(14): 4509-4517. HUANG F Z, DING T, LI X K, et al. Species diversity for various associations along an altitudinal gradient in the karst seasonal rainforest in Nonggang[J]. Acta Ecologica Sinica, 2016, 36(14): 4509-4517. |
| [13] |
俞筱押, 余瑞, 黄娟, 等. 贵州茂兰喀斯特森林四药门花群落优势种群生态位特征[J]. 生态学杂志, 2017, 36(12): 3470-3478. YU X Y, YU R, HUANG J, et al. Niche characteristics of dominant plant populations of Loropetalum subcordatum community in Maolan karst forest, Guizhou[J]. Chinese Journal of Ecology, 2017, 36(12): 3470-3478. |
| [14] |
王斌, 黄俞淞, 李先琨, 等. 弄岗北热带喀斯特季节性雨林15 ha监测样地的树种组成与空间分布[J]. 生物多样性, 2014, 22(2): 141-156. WANG B, HUANG Y S, LI X K, et al. Species composition and spatial distribution of a 15 ha northern tropical karst seasonal rain forest dynamics study plot in Nonggang, Guangxi, southern China[J]. Biodiversity Science, 2014, 22(2): 141-156. |
| [15] |
黄甫昭, 王斌, 丁涛, 等. 弄岗北热带喀斯特季节性雨林群丛数量分类及与环境的关系[J]. 生物多样性, 2014, 22(2): 157-166. HUANG F Z, WANG B, DING T, et al. Numerical classification of associations in a northern tropical karst seasonal rain forest and the relationships of these associations with environmental factors[J]. Biodiversity Science, 2014, 22(2): 157-166. |
| [16] |
梁畴芬, 梁健英, 刘兰芳, 等. 弄岗自然保护区植物区系考察报告[J]. 广西植物, 1988, 8(增刊1): 83-184. LIANG C F, LIANG J Y, LIU L F, et al. A report on the floristic survey on the Nonggang Natural Reserve[J]. Guihaia, 1988, 8(Suppl.1): 83-184. |
| [17] |
黄俞淞, 吴望辉, 蒋日红, 等. 广西弄岗国家级自然保护区植物物种多样性初步研究[J]. 广西植物, 2013, 33(3): 346-355. HUANG Y S, WU W H, JIANG R H, et al. Primary study on species diversity of plant in Nonggang National Natural Reserve of Guangxi[J]. Guihaia, 2013, 33(3): 346-355. DOI:10.3969/j.issn.1000-3142.2013.03.011 |
| [18] |
苏宗明. 广西(山王丌)弄岗石灰岩山森林类型的分类问题[J]. 广西植物, 1981, 1(2): 7-10. SU Z M. Classification of limestone forest in Nonggang, Guangxi[J]. Guihaia, 1981, 1(2): 7-10. |
| [19] |
CONDIT R. Research in large, long-term tropical forest plots[J]. Trends in Ecology and Evolution, 1995, 10(1): 18-23. DOI:10.1016/S0169-5347(00)88955-7 |
| [20] |
高贤明, 陈灵芝. 北京山区辽东栎(Quercus liaotungensis)群落物种多样性的研究[J]. 植物生态学报, 1998, 22(1): 23-32. GAO X M, CHEN L Z. Studies on the species diversity of Quercus liaotungensis communities in Beijing Mountains[J]. Acta Phytoecologica Sinica, 1998, 22(1): 23-32. DOI:10.3321/j.issn:1005-264X.1998.01.003 |
| [21] |
王刚. 植物群落中生态位重迭的计测[J]. 植物生态学与地植物学丛刊, 1984, 8(4): 329-335. WANG G. On the measurement of niche overlap in plant communities[J]. Acta Phytoecologioaet Geobotanica Sinica, 1984, 8(4): 329-335. |
| [22] |
PIANKA E R. The structure of lizard communities[J]. Annual Review of Ecology & Systematics, 1973, 4: 53-74. |
| [23] |
国际生物多样性计划中国委员会. 中国生物多样性保护与研究进展X[M]. 北京: 气象出版社, 2014. CNC-DⅣERSITAS. Advances in biodiversity conservation and research in China X[M]. Beijing: China Meteorological Publishing House, 2014. |
| [24] |
刘润红, 常斌, 荣春艳, 等. 漓江河岸带枫杨群落主要木本植物种群生态位[J]. 应用生态学报, 2018, 29(12). LIU R H, CHANG B, RONG C Y, et al. Niche of main woody plant populations of Pterocarya stenoptera community in riparian zone of Lijiang River, China[J]. Chinese Journal of Applied Ecology, 2018, 29(12). DOI:10.13287/j.1001-9332.201812.001 |
| [25] |
王斌, 黄俞淞, 李先琨, 等. 广西弄岗喀斯特季节性雨林——树种及其分布格局[M]. 北京: 中国林业出版社, 2016. WANG B, HUANG Y S, LI X K, et al. Guangxi Nonggang karst seasonal rain forest: Tree species and their distribution patterns[M]. Beijing: China Forestry Publishing House, 2016. |
| [26] |
史作民, 程瑞梅, 刘世荣. 宝天曼落叶阔叶林种群生态位特征[J]. 应用生态学报, 1999, 10(3): 265-269. SHI Z M, CHENG R M, LIU S R. Niche characteristics of plant populations in deciduous broad-leaved forest in Baotianman[J]. Chinese Journal of Applied Ecology, 1999, 10(3): 265-269. DOI:10.3321/j.issn:1001-9332.1999.03.003 |
| [27] |
张忠华, 梁士楚, 胡刚. 桂林岩溶石山阴香群落主要种群生态位研究[J]. 林业科学研究, 2009, 22(1): 63-68. ZHANG Z H, LIANG S C, HU G. Niche characteristics of dominant populations in Cinnamomum burmannii community on karst hills of Guilin[J]. Forest Research, 2009, 22(1): 63-68. DOI:10.3321/j.issn:1001-1498.2009.01.011 |
| [28] |
蒋忠诚, 罗为群, 童立强, 等. 21世纪西南岩溶石漠化演变特点及影响因素[J]. 中国岩溶, 2016, 35(5): 461-468. JIANG Z C, LUO W Q, TONG L Q, et al. Evolution features of rocky desertification and influence factors in karst areas of southwest China in the 21st century[J]. Carsologica Sinica, 2016, 35(5): 461-468. |
| [29] |
李先琨, 何成新, 唐建生, 等. 广西岩溶山地生态系统特征与恢复重建[J]. 广西科学, 2008, 15(1): 80-86, 91. LI X K, HE C X, TANG J S, et al. Evolution and ecological processes of karst ecosystem of Guangxi[J]. Guangxi Sciences, 2008, 15(1): 80-86, 91. DOI:10.3969/j.issn.1005-9164.2008.01.023 |
| [30] |
袁正科. 拟生造林法的提出与应用实例[J]. 湖南林业科技, 1996, 23(3): 1-6. YUAN Z K. Introduction of near natural afforestation and application examples[J]. Journal of Hunan Forestry Science and Technology, 1996, 23(3): 1-6. |
| [31] |
丛沛桐, 颜延芬, 周福军, 等. 东北羊草群落种群生态位重叠关系研究[J]. 植物研究, 1999, 19(2): 212-219. CONG P T, YAN Y F, ZHOU F J, et al. The study on ecological niche overlap relations of several populations on Leymus community in northeast plain[J]. Bulletin of Botanical Research, 1999, 19(2): 212-219. DOI:10.3969/j.issn.2095-0845.1999.02.014 |
| [32] |
陈俊华, 刘兴良, 何飞, 等. 卧龙巴朗山川滇高山栎灌丛主要木本植物种群生态位特征[J]. 林业科学, 2010, 46(3): 22-28. CHEN J H, LIU X L, HE F, et al. Niche characteristics of dominant woody populations in Quercus aquifoliodes shrub community in Balangshan Mountain in Wolong Nature Reserve[J]. Scientia Silvae Sinicae, 2010, 46(3): 22-28. |
| [33] |
周亚琦, 官凤英, 范少辉, 等. 天宝岩竹阔混交林毛竹及其伴生树种生态位的研究[J]. 北京林业大学学报, 2017, 39(7): 46-53. ZHOU Y Q, GUAN F Y, FAN S H, et al. Niche characteristics of Phyllostachys edulis and its associated tree species in Tianbaoyan bamboo and broadleaved mixed forest[J]. Journal of Beijing Forestry University, 2017, 39(7): 46-53. |
| [34] |
钱逸凡, 伊力塔, 胡军飞, 等. 普陀山主要植物种生态位特征[J]. 生态学杂志, 2012, 31(3): 561-568. QIAN Y F, YI L T, HU J F, et al. Niche characteristics of main plant species in Putuo Mountain, Zhejiang Province of East China[J]. Chinese Journal of Ecology, 2012, 31(3): 561-568. |
| [35] |
李显森, 于振海, 孙珊, 等. 长江口及其毗邻海域鱼类群落优势种的生态位宽度与重叠[J]. 应用生态学报, 2013, 24(8): 2353-2359. LI X S, YU Z H, SUN S, et al. Ecological niche breadth and niche overlap of dominant species of fish assemblage in Yangtze River estuary and its adjacent waters[J]. Chinese Journal of Applied Ecology, 2013, 24(8): 2353-2359. |
| [36] |
陈波, 周兴民. 3种嵩草群落中若干植物种的生态位宽度与重叠分析[J]. 植物生态学报, 1995, 19(2): 158-169. CHEN B, ZHOU X M. Analyses of niche breadths and overlaps of several plant species in three Kobresia communities of an alpine meadow[J]. Acta Phytoecologica Sinica, 1995, 19(2): 158-169. DOI:10.3321/j.issn:1005-264X.1995.02.007 |
| [37] |
刘屹, 邓竣尹, 谢天资, 等. 生态位研究进展及干旱河谷生态位研究重点[J]. 四川林业科技, 2015, 36(5): 16-20. LIU Y, DENG J Y, XIE T Z, et al. Advances in niche research and niche research priorities in dry valley areas[J]. Journal of Sichuan Forestry Science and Technology, 2015, 36(5): 16-20. DOI:10.3969/j.issn.1003-5508.2015.05.004 |
| [38] |
黄丽君, 卢艳春, 徐冬英, 等. 苹婆的栽培现状及发展对策[J]. 中国热带农业, 2014(3): 36-37. HUANG L J, LU Y C, XU D Y, et al. The cultivation status and development strategy of Sterculia nobilis Smith[J]. China Tropical Agriculture, 2014(3): 36-37. DOI:10.3969/j.issn.1673-0658.2014.03.015 |
| [39] |
张俊杰, 柴胜丰, 韦霄, 等. 珍稀濒危植物金丝李种子的萌发特性[J]. 林业科学, 2018, 54(4): 174-185. ZHANG J J, CHAI S F, WEI X, et al. Germination characteristics of the seed of a rare and endangered plant, Garcinia paucinervis[J]. Scientia Silvae Sinicae, 2018, 54(4): 174-185. |
| [40] |
傅立国. 中国植物红皮书:稀有濒危植物[M]. 北京: 科学出版社, 1991. FU L G. China plant red data book:Rare and endangered plants[M]. Beijing: Science Press, 1991. |
| [41] |
张俊杰, 柴胜丰, 吕仕洪, 等. 珍稀濒危植物金丝李的生境特征及致濒原因分析[J]. 生态环境学报, 2017, 26(4): 582-589. ZHANG J J, CHAI S F, LV S H, et al. The habitat characteristics and analysis on endangering factors of rare and endangered plant Garcinia paucinervis[J]. Ecology and Environmental Sciences, 2017, 26(4): 582-589. |
| [42] |
赵永华, 雷瑞德, 何兴元, 等. 秦岭锐齿栎林种群生态位特征研究[J]. 应用生态学报, 2004, 15(6): 913-918. ZHAO Y H, LEI R D, HE X Y, et al. Niche characteristics of plant population in Quercus aliena var.Acuteserrata stands in Qinling Mountain[J]. Chinese Journal of Applied Ecology, 2004, 15(6): 913-918. DOI:10.3321/j.issn:1001-9332.2004.06.001 |
| [43] |
陈玉凯, 杨琦, 莫燕妮, 等. 海南岛霸王岭国家重点保护植物的生态位研究[J]. 植物生态学报, 2014, 38(6): 576-584. CHEN Y K, YANG Q, MO Y N, et al. A study on the niches of the state's key protected plants in Bawangling, Hainan Island[J]. Chinese Journal of Plant Ecology, 2014, 38(6): 576-584. |
| [44] |
何忠伟, 胡仁传, 黄日波, 等. 广西银杉林主要树种种群生态位分析[J]. 林业科学研究, 2012, 25(6): 761-766. HE Z W, HU R C, HUANG R B, et al. Niche of the major tree species populations in Cathaya argyrophylla forest in Guangxi[J]. Forest Research, 2012, 25(6): 761-766. DOI:10.3969/j.issn.1001-1498.2012.06.014 |